P03 - Supramolecular organization of ATP synthase and protonic energy coupling
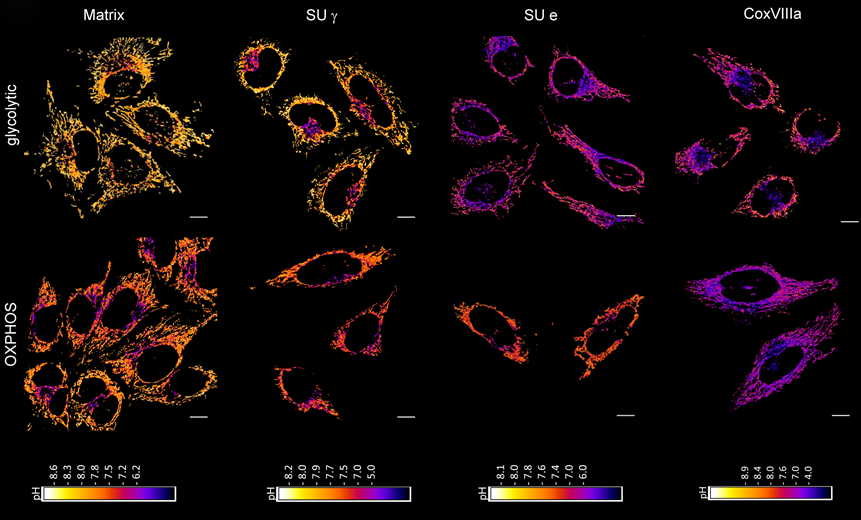
Mitochondrial ATP synthase is an enzyme pivotal for mitochondrial and cellular physiology. Energy derived from oxidative metabolism is stored in a proton-motive force (PMF) across the inner membrane of the organelle. The ATP synthase (CV) harness the PMF to generate ATP but also contributes to maintain the PMF under unfavourable conditions. Human ATP synthase is a complex of 29 subunits (including the regulatory protein IF1). The rotary machine consists of a membrane part (F0) and membrane-extrinsic (F1) part linked by central and peripheral stalks. Mitochondrial F1FO ATP synthase is organized in dimers that form rows along the rims of cristae. This specific supramolecular structure also contributes to the formation and stabilization of the specific cristae architecture, next to the cristae-junction related protein complexes OPA1 and the MICOS complex. Functional and physical interaction between ATP synthase and OPA1, respectively MICOS compounds were reported, suggesting a crosstalk between the cristae-shaping proteins. The dimerization of ATP synthase is promoted by subunits e and g, while the subunit DAPIT/USMG5 is likely involved in dimer-dimer stabilization. Likewise, the inhibitory factor IF1 also promotes dimer-dimer interaction in addition to its ATP synthase activity-regulating function. We recently showed that also the spatiotemporal organization of ATP synthase and its mobility in cristae membranes is determined by IF1. In the upcoming funding period, we aim to better understand the relationship between the supramolecular organization of ATP synthase and its enzyme activity. To this end, we will manipulate the oligomerization of ATP synthase via changing concentrations of the DAPIT- and IF1 subunits. The effects of decreased CV oligomerization on enzyme activity will be measured in living cells using fluorescent biosensors for pH, m, and ATP. We hypothesize that disruption of oligomeric structure leads to enzyme instability and a shift toward ATP hydrolysis, likely also involving ultrastructural changes of cristae. This could be counterbalanced by upregulation of IF1. In collaboration with P02, we will study the crosstalk of DAPIT/ATP synthase with the MICOS complex, specifically the Mic10 subunit. Our cell lines will be made available for additional studies within the consortium (P05, P06, P07) to further investigate the crosstalk between supramolecular ATP organization and MICOS.